Thèse présentée au Conservatoire National des Arts et Métiers
pour obtenir le diplôme d’Ingénieur
par
A. SDIKA
CONTRIBUTION A L’ETUDE DES CIRES DE VIGNES EN RELATION AVEC L’AGE ET LA RESISTANCE AU MILDIOU
soutenue en décembre 1964
INTRODUCTION
Le but du travail que nous avons entrepris a trait à l'étude comparé des fractions cireuses de deux espèces de vignes - l'une résistante et l'autre sensible au Mildiou - envisagée des poins de vue pondéral et qualitatif.
Ce travail se situe dans le cadre des recherches normalement menées à l'IRCHA, d'une part sur les causes et les conditions agronomiques de la résistance des plantes à certaines maladies parasitaires, d'autre part sur l'étude et les possibilités de synthèse de substances fongistatiques et fongicides.
Les grandes lignes de cette étude ont consisté d'abord en la préparation dans des conditions de reproductibilité satisfaisante, des fractions aux substances pures destinées aux essais physiologiques, puis en la détermination et en la comparaison de leurs constantes physico-chimiques, de leurs caractères analytiques, et de leur importance respective dans le mélange complexe qu'est la cire.
Cette comparaison a été faite en fonction de l'espèce et de l'âge de la feuille.
Elle peut faire ressortir entre les métabolismes des deux espèces des différences qu'il serait éventuellement possible de modifier par des voies artificielles.
Un ou plusieurs des facteurs de la résistance au Mildiou sont ainsi susceptibles d'apparaître.
Bien que bornant notre étude aux constituants de la cire cuticulaire, nous n'exclurons pas, le cas échéant, celle de substances plus rarement rencontrées dans les cires.
Très rapidement s'est posé à nous le problème d'une définition précise du matériel à étudier, ceci autant pour le délimiter que pour baser nos comparaisons et les rendre valables.
La cire a en effet une définition imprécise.
Il convient, dès l'abord, de la replacer dans un contexte fonctionnel, c'est-à-dire celui d'un ensemble de substances formant revêtement et possédant la propriété de s'opposer, soit par ses propriétés physico-mécaniques, soit par la nature chimique de ses constituants -ou par les deux- à la germination puis à l'infection par le parasite.
Plusieurs auteurs ont tenté de définir les cires mais avec une rigueur relative, en se basant plus ou moins sur leur ressemblance avec la cire d'abeilles et leur composition chimique.
Ainsi LEO IVANOVSKY les définit par tout un ensemble de caractères physiques et pratiques (dont celui de pouvoir façonner des chandelles allant de pair avec une composition chimique comportant des hydrocarbures, des esters, des acides et alcools lourds et des composés aromatiques.
POLONWSKY les considère comme un mélange de cérides -esters d'un acide gras et d'un monoalcool aliphatique (cérides vrais - ou cyclique (stérides) si l'alcool est un stérol).
Au sens large, les cires renferment aussi des hydrocarbures, acides, cétones et alcools de haut poids moléculaire, à propriété proches de ceux des cérides.
MAZLIAK reconnaît que dans le langage commun on appelle parfois cires des mélanges de glycérides, alors que des huiles (huile de Spermaceti) par leur composition, devraient être rangées parmi les cires.
Si l'on se réfère systématiquement à la composition des mélanges naturels appelés cires, on y trouve des substances très variées, et parfois à des taux importants : La Cire de Cochinchine, provenant du fruit Ironigia basteri et I. gabonensis renferme 90% de triglycérides.
La cire d'Ucuhuba (Myristica surinamensis et Virola surinamensis) contient 65 à 76% de triglycérides et 7% de résines.
Il en est de même de la cire d'arbre de chine (Sapicum sebiferum).
Beaucoup de ces cires comportent un "soluble alcool" formé de résines, de stérols, d'acides, de matières volatiles (jusqu'à 14% dans la cire de Candellila) et 20% de stéroïdes (phytostérols et isovalérates d'amyrénol) dans la cire des feuilles de diverses espèces de Ceanothus.
La cire de coton, semi raffinée, renferme 12% des mêmes impuretés etc.
On remarque que dans la généralités des cas, ces impuretés sont solubles dans les cires, les huiles et les solvants des lipides, qu'elles sont hydrophobes, et constituent souvent avec la ligne cellulose la paroi protectrice des cellules.
Il est possible de restreindre la définition de la cire en la ramenant à celle du mélange des constituants d'une chaîne de précurseurs et d'intermédiaires biogénétiques.
L'un des schémas proposé pour la biosynthèse des constituants des cires et celui de Chibnall et Pipo en 1934.
"II rend compte de la présence d'une série d'acides gras et d'alcools primaires à nombre pair d'atomes de carbone aussi bien que d'alcools secondaires, de cétones et de paraffines un nombre impair d'atomes de carbone.
On part d'un précurseur acide gras insaturé à nombre pair d'atomes de carbone et dont le nombre et la placides doubles liaisons sont discutés.
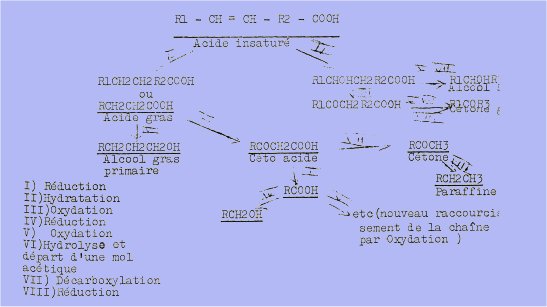
Les esters (cérides, stérides) se formeraient selon WARTH par un processus simple d’estérification.
Quant aux hydrocarbures ils proviennent de la décarboxylation de ces esters :
RCOOCH2R’ ——> CH2R’ + C02.
Selon WANLESS. ils dériveraient des acides mêmes :
RC2-CH2-CH2-COOH ——> RCH2-CO-CH2-COOH ——> RCH2-CO-CH3 ——> RCH2-CH2-CH3
Par contre, selon MAZLIAK, les paraffines résulteraient d'un métabolisme indépendant de celui des acides gras.
Cet auteur estime, en se basant sur des expériences réalisées avec des molécules marquées que les acides gras saturés et insaturés des cires sont synthétisés d'abord par condensation de fragments acétiques.
Les alcools gras, les diols ou les hydroxyacides pourraient dériver des acides homologues par réduction ou oxygénation.
Les composés cycliques seraient synthétisés à partir de fragments acétiques et condensation de précurseurs isopréniques.
Le nombre des atomes de carbone des constituants des cires varie entre 12 et 32. Les cérides de plus bas poids moléculaire connus sont : le laurate de cétyle (C12-C16) PF 41° PM 425, et le Myristate de Myristile (C14-C14) PF 38,5. PM 425.
En ce qui concerne les feuilles de vignes, l'examen de la littérature a montré que, jusqu'à présent à notre connaissance aucun travail de cette nature sur leur cire n'avait été entrepris.
Les perspectives ouvertes par la découverte de l'existence de propriétés fongistatiques chez les cires de feuilles de pommier et chez celles d'autres plantes accentuent l'intérêt de cette recherche.
LES CAUSES DE LA RESISTANCE
Parmi les champignons qui attaquent les vitacés, l'un des plus importants est le Plasmopora, Siphonycete Oomycète représenté par plusieurs espèces dont le Plasmopora Viticola qui parasite les vignes des régions européennes (Vitis Vinifera).
Au cours de nos recherches bibliographiques, nous avons relevé plusieurs observations de biologistes présentant une relation avec notre travail.
D. BOUBALS remarque que les mucilages apportés par les conidies rendent le limbe plus mouillable, facilitant ainsi l'infection ; l'attraction elle même est due à un phénomène de chimiotactisme.
Selon GAUMN, il existe un seuil numérique d'infection soumis à plusieurs facteurs dont le temps de persistance de l'eau sur les feuilles, temps suffisant pour que germent les sporanges et que les Zoopores se fixent aux stomates et mettent un tube germinatif.
Le mycélium s'étend plus vite dans les tissus jeunes que dans ceux, plus âgés, qui possèdent des tissus "barricades".
Les sporanges doivent séjourner au minimum 1H10 pour que s'établisse l'infection qui est, d'autre part, favorisée par l'humidité de l'air.
Les sporanges eux-mêmes sont peu influencés par le pH du milieu.
MYSTERAKIS, en 1943 signale qu'il a pu en faire germer dans du jus de feuilles de V. Riparia à pH 2-3.
Cependant, l’eau de pluie en contact avec les feuilles a toujours une action alcaline (Diffusion des sels en cours de transpiration).
Dans nos laboratoires d'essais physiologiques, il a été fréquemment observé que. pour une variété non résistante, le degré de l'attaque varie considérablement avec l'âge de la feuille.
A la lignification des pétioles, la résistance augmente alors que la feuille jeune d'un clône est contaminable in vitro.
Si on lave la pellicule cireuse par des solvants de lipides ne lésant pas le parenchyme cellulaire d'une feuille résistante, la contamination devient réalisable.
Il a été d'autre part observé un phénomène lié à la nature de la cire ou à une orientation des groupements hydrophiles :
Une jeune feuille cultivée en serre pas mouillable, contrairement à la feuille de même âge cultivée à l'air libre, et de l'eau pulvérisée sur la première roule en gouttelettes.
Mais 48 h plus tard. une nouvelle pulvérisation montre l'apparition d'une certaine mouillabilité.
Enfin, il a été constaté que les fongicides organiques à base de sel de cuivre et de zinc montrent une efficacité amoindrie chez la feuille adulte après l'aoûtement.
Une rapide revue de bibliographie, portant sur les trente dernière années va nous permettre de résumer les travaux et les hypothèses de divers chercheurs, essentiellement des biologistes :
En 1933, B. HUSFELD étudiant la transmission héréditaire des causes de la résistance attribue celle-ci à la formation des nécroses dans les tissus des espèces résistantes en constituant une barrière à l'égard du développement du mycélium.
Il n'explique pas le mécanisme de leur formation et de leur intervention. En outre, les substances du suc cellulaire gêneraient le développement du mycélium dans les tissus lésés.
Il conclut à l'hérédité polyfactorielle des causes qui, chez les espèces résistantes, provoquent ces nécroses.
En France, NYSTERAKIS en 1943 élimine de nombreuses causes possibles: concentration du suc cellulaire, acidité totale et pH du jus des feuilles, teneur en eau, teneurs en azote, potasse P04, calcium, aminoacides, amidon, sucres.
De plus le jus des feuilles résistantes n'est pas toxique pour le Plasmopora.
Enfin il n'a pu mettre en évider: la production, dans les tissus cicatriciels de V. VINIFERA de composés chimiques capables d'enrayer l'extension du parasite ou de neutraliser ses éventuelles sécrétions toxiques.
Il explique finalement la résistance des vignes américaines et de leurs hybrides par la forte sensibilité des tissus dont la réaction provoquerait la mort du mycélium.
G. MOREL en 1948 remarqua que les tissus des vignes sensibles issus des repiquages résistent mieux à la prolifération que ceux des cals primitifs.
COUTINH0 en 1950 affirme l'existence de degré, de résistance à l’intérieur de V. VINIFERA L.
L.C. PIOTH en 1957 a recherché les différences existant entre plantes sensibles et résistantes au point de vue anatomique et composition chimique étudiée par chromatographie sur papier du jus des feuilles : il a observé chez les feuilles parasitées une concentration du mycélium au niveau des cellules renfermant des cristaux d'oxalate de calcium.
Les cellules sont nombreuses chez les sensibles à toute époque du cycle végétatif, alors qu'elles se trouvent en nombre inférieur chez les variétés résistantes V. Biparia et V. Rupestris.
Mais il remarque des exceptions.
Cet auteur a aussi mis en évidence des composés indoliques chez les feuilles de vignes sensibles et qui n’apparaissent pas dans les variétés très résistantes.
Chez les vignes moins résistantes ces composés n'apparaissent qu'à partir du ralentissement de la croissance. Mais il n'y a pas de parallélisme entre sensibilité et concentration.
L'acide oxalique et les composés indoliques (qui ont le caractère de substances de croissance) joueraient donc un grand rôle dans les phénomènes de sensibilité au Mildiou.
Cependant si des plantes résistantes traitées par les acides indolylacétique et indolybutyrique deviennent plus sensibles, paradoxalement des plantes sensibles traitées par les mêmes substances, en deviennent plus résistantes.
En faisant entrer dans la composition d'un milieu de culture l'acide indolacétique ainsi que l'asparagine, l'aneurine et l'acide oxalique, il aurait obtenu la formation de sporangiophores.
DENIS BOUBALS estime accessoire le rôle de la couche cireuse et de la villosité et qui est d'éviter de mouiller limbe, et, par là, les zoospores d'atteindre les stomates (résistance à l'infection proprement dite).
C'est la résistance à l'expansion du champignon chez l'hôte qui est la plus importante (tissus barricades chez les espèces sensibles).
Chez les espèces résistantes, ce sont les phénomènes nécrotiques qui s'y opposent, mais "Tout se passe comme si lorsqu'un facteur fait défaut ou se trouve on trop faible quantité, aucun développement du mycélium ne se produit après la pénétration des zoospores dans les stomates..."
Il démontre ainsi que l'hypersensibilité de l'hôte, qui se manifeste par les nécroses somatiques et périsomatiques et les dessications internervales du limbe, est secondaire.
Nous observerons que les hypothèses et conclusions de travaux de ces auteurs n'excluent pas, chez les feuilles résistantes, l'intervention de facteurs biochimiques d'inhibition des zoospores du Mildiou ou - à un stade ultérieur - de la dessication du mycélium.
Elles n'excluent pas également la possibilité d'existence, chez les espèces sensibles, de nouveaux composés - spécifiques ou non - du développement du Mildiou.
En outre les travaux sur les causes possibles de la résistances n'ont pas porté sur toutes les classes de composés existant dans la feuille en particulier les lipides, les résines, les stéroïdes.
CONCLUSION
Nous allons, au terme de ce travail, tenter de. dégager et d'interpréter les résultats dans leur ensemble.
Ces résultats obtenus à partir de deux échantillons, devront, pour présenter un caractère général, être complétés par des études ultérieures similaires sur d’autres espèces de feuilles de vigne.
Ils seront considérés de quatre points de vue : l) Analyse de la fraction cireuse. 2) Etude du métabolisme de certains constituants, ce qui représente l'aspect fondamental de nos recherches. 3) Apports et modifications de techniques nécessités par les particularités de notre matériel d'étude. 4) Hypothèses de travail complémentaires quant aux causes de la résistance an mildiou.
l) Analyse de la fraction cireuse :
a)Au point de vue qualitatif, nous avons pu isoler et identifier -pour la première fois, semble t'il - le n-nonacosane et l'alcool cérylique dans les feuilles de vigne.
Ces deux corps semblent, tout au moins chez les jeunes feuilles, les seuls de leur série constituant la cire vraie.
Nous avons en outre isolé et identifié chez Noah2 le stéarate de céryle, et isolé sans l’identifier, un ester insaturé dans CFIII.
Il est probable que ces deux esters existent, le premier dans Noah1 et CFI, et le second dans CFII et CFI.
Il semble intéressant, au point de vue de la biogenèse de ces composés, de noter que cet ester insaturé ne se trouve pas dans Noah1, ce qui impliquerait soit un système différent d’oxydoréduction enzymatique, soit une différence d'activité, pour une cause à déceler, de réductases et d’estérases.
Cette hypothèse supputerait également sur la prédominance de l'acide linoleïque chez les chasselas et de l'oleïque chez les Noah.
Ce qui distingue les deux cires vraies, c'est également la présence des stérides chez les seuls Noah.
Nous remarquerons cependant que ces stérides ont pu se trouver en faible proportion dans les cires de Chasselas, mais se trouver éliminés à la suite, des recristallisations.
L’un des stérols estérifiés est probablement Ie sitostérol.
Un acide triterpénique, l’acide oléanolique a également été identifié à l'état libre. Il semble être le seul de cette classe de composés.
Il existe chez les deux espèces de feuilles et ne paraît donc pas jouer un rôle quelconque dans les phénomènes de résistance.
Cependant, nous n'en avons pas déterminé la teneur, ni comparé les formes sous lesquelles il se trouve.
L’ergostérol ne parait pas non plus jouer un rôle dans les phénoménes de résistance, sa présence étant, au même titre, douteuse dans les huit cires étudiées.
Ont été d’autre part identifiés sans isolement des acides déjà découverts dans la vigne : Les acides oleïque, linoleïque et palmitique dans les Chasselas et les acides oleïque et palmitique dans les Noah.
L'acide stéarique a été identifié dans la cire vraie, sous forme uniquement combinée.
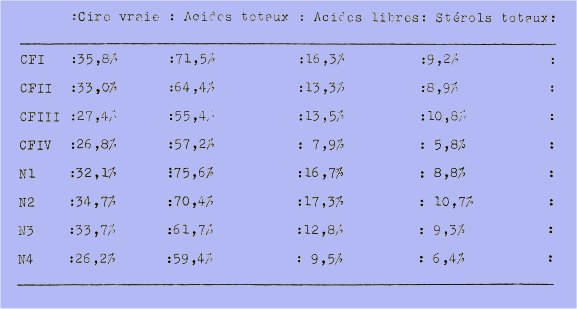
b) Au point de vue quantitatif : Nous résumerons dans un tableau les résultats rapportés à la cire résineuse :
2) Etude comparée de l'évolution des constituants cireux avec l'âge.
En ce qui concerne, la cire totale, sa teneur ne subit pas de modifications significatives avec l'âge et ce, quelle que soit la cire considérée dans ses divers degrés de purification.
On note, toutefois un accroissement général des cires des deux espèces de feuilles à la quatrième récolte, bien que la teneur en cire vraie n'en soit pas affectée.
Pour une comparaison valable des teneurs, on doit cependant reconnaître qu'il y aurait eu autant de raisons de rapporter les poids de cire à ceux des feuilles sèches, qu'à la surface de ces dernières, puisque la protection des feuilles dépend en grande partie de l'épaisseur, de l’étalement et des propriétés mécaniques de la couche cireuse.
Nous avons déjà noté, dans la mesure des indices, la valeur remarquable de l'indice d'iode des jeunes feuilles de Chasselas, puis son abaissement rapide.
Interprétées dans le sens d'une intervention enzymatique, les évolutions et les différences dans l'insaturation traduiraient une activité plus stable et plus marquée des réductases du Noah.
Les techniques chromatographiques nous ont montré la disparition, dans les cires de Chasselas, de l'acide oléïque des feuilles en voie de croissance, et la présence permanente de l'acide linoléïque.
Les acides libres désignés tant par l'indice d'acide que par la teneur diminuent avec l'âge alors que les esters augmentent légèrement.
Le rapport des deux indices, traduit sur graphique, montre une similitude d'évolution notable.
Il en est de même des deux couples de courbes indices iode-hydroxyle, bien que dans une moindre mesure.
3) Apports et modifications techniques :
Au cours de notre travail, nous avons dû mettre au point ou modifier un certain nombre de techniques de façon à les rendre applicables à nos cires et ce, dans la perspective d'un meilleur rendement ou d'une reproductibilité améliorée.
Ainsi, le choix du mode d'extraction de la fraction lipido-cireuse totale, la purification des cires de vigne par dépigmentation, la récupération quantitative des acides libres avec séparation sélective des acides supérieurs.
Nous avons été amené à déterminer l'indice d’acide par un dosage en retour, et l'indice d’hydroxyle par une technique d'acétylation des graisses qui s'est révélée seule reproductible.
Dans le dosage des stérols de feuilles de vigne, nous avons été amené à adapter la technique de WALL et KELLEY à nos extraits benzéniques de cire résineuse, puis à juger de la valeur de la méthode d’ISSIDORIDES dans son application à la récupération des stérols de nos feuilles.
Nous avons vu qu'elle est satisfaisante.
Enfin, dans l'isolement des esters de Noah2 et de CFIII, nous avons combiné plusieurs techniques de chromatographie sur colonne pour parvenir à ce résultat.
4) Hypothèses de travail :
Notre travail nous semble ouvrir la voie à plusieurs hypothèses qui peuvent se révéler complémentaires :
En faveur d'une théorie nutritionnelle du mildiou (parasite strict) il semblerait qu'un taux minima de stérols libres et d'acide linoléïque soit nécessaire.
Ce dernier, entrant avec l'acide linolénique (autre acide polyinsaturé) dans le complexe vitaminique F il serait d'un certain intérêt d’en. étudier le rôle dans la croissance du parasite.
Cependant, nous n'avons pas -bien que cet acide ait été déjà isolé de la vigne- observé d'acide linolénique.
Il convient, d’autre part d'éliminer les acides triterpéniques libres en tant que substances fongistatiques ou dynamisantes, mais le rôle de la saponine même reste à étudier.
Nous croyons peu probable l'existence d'acides fongistatiques dans la fraction A (acides gras supérieurs), mais les fractions B1 et surtout B2 des deux dernières récoltes, renfermant des substances polyphénoliques, pourraient se révéler intéressantes dans les essais physiologiques.
Nous avons vu qu'ils apparaissent dans la fraction cireuse de la feuille adulte, à partir de la troisième récolte.
Enfin en ce qui concerne la différence de qualité des barrières cireuses des deux espèces de feuilles, nous avons déjà noté qu'elle paraît uniquement due à l’insaturation et à la fluidité de celle de CFI par rapport à Noahl.
MOTS CLEFS : âge / alcool / cause / céride / champignon / chasselas / chimique / cire / comparaison / constituant / croissance / ester / étude / facteur / feuille / fongicide / huile / hydrocarbure / hypothèse / infection / limbe / linoleïque / mildiou / mycélium / nécroses / noah / oleïque / paraffine / parasite / plantes / propriété / résines / résistance / sensible / stéride / stérol / stomate / substance / technique / tissus / vigne / zoospore