ThŤse prťsentťe ŗ la facultť des sciences de líUniversitť de Paris
pour obtenir le grade de Docteur es sciences-physiques
par
Fernando DI JESO
OPHELINE, PHOSPHO-OPHELINE, OPHELINE-PHOSPHOTRANSFERASE
soutenue LE 4 JUIN 1965
devant la commission díexamen :
E. Lederer, prťsident
F. Chapeville, Van Thoaiexaminateurs
INTRODUCTION
Les phosphagŤnes constituent un chapitre de biochimie comparťe qui a beaucoup ťvoluť ces derniŤres annťes.
Il s'est ouvert, il y a trente cinq ans par les recherches de Fiske et Subbarow (1927, 1929), d'Eggleton et Eggleton (1927, 1928, 1929') et de Meyerhof et Lohmann (l928, 1928) qui conduisirent ŗ la dťcouverte de la phosphocrťatine (Fiske et Subbarow, 1929) et de la phosphoarginine (Meyerhof et Lohmann, 1928).
La synthŤse de la premiŤre de ces deux substances a ťtť rťalisťe beaucoup plus tard. par Zeile et Fawaz (1958). Ennor et Stocken (1948). Pradel. Thiem. Pin et Thoai (1959) ; celle de la seconde ne l'a ťtť rťalisťe qu'en 1962 par Thiem, Thoai et Roche, et par Cramer, Schieffele et Vollmar (1962), selon deux mťthodes diffťrentes.
La phosphoglycocyamine et la phosphotaurocyamine ont ťtť dťcouvertes, il y a dix ans. par Thoai et collaborateurs (1955, 1954), isolťes et synthťtisťes par Thoai et Thiem en 1957.
La phospholombricine a ťtť dťcouverte en 1955 par Thoai et Robin (1954) et synthťtisťe en I960 par Beatty, Ennor et Magrath.
La phosphohypotaurocyamine a ťtť identifiťe par Robin et Thoai en 1962 ; elle nía pas encore ťtť synthťtisťe, mais elle a ťtť obtenue ŗ un haut degrť de puretť par transphosphorylation enzymatique en 1962 par Thoai, Robin et Pradel.
AprŤs les premiŤres ťtudes poursuivies sur la rťpartition de la phosphocrťatine, de la phosphoarginine et des deux bases guanidiques libres dans le rŤgne animal, il a ťtť gťnťralement admis que la rťpartition de ces dťrivťs guanidiques suit un ordre phylogťnťtique (Needham, Needham, Baldwin et Yudkin, 1952).
En effet, on avait retrouvť la crťatine (Hunier, 1928) et la phosphocrťatine (Eggleton et Eggleton, 1928) chez les Vertťbrťs et non chez les Invertťbrťs, dont les muscles ne paraissaient renfermer que de l'arginine et de la phosphoarginine (Baldwin et Needham, 1954 ; Eggleton et Eggleton, 1928 ; Meyerhof, 1928, Needham, Needham, Baldwin et Yudkin, 1952 ; Schutze, 1952).
Plus tard, on a mis en ťvidence la phosphocrťatine. mais non la phosphoarginine. chez des Protochordťs ; de plus, ces deux phosphagŤnes coexistent chez des Echinoidťs.
Ces donnťes semblaient confirmer les thťories de l'ťvolution, selon lesquelles une transition biochimique entre Vertťbrťs et Invertťbrťs ťtait observťe au mÍme niveau de la classification que celui auquel se manifeste une transition morphologique entre ces deux groupes zoologiques (Baldwin et Yudkin. 1950).
Depuis dix ans un nombre important de recherches. poursuivies surtout sur les Annťlides, mais aussi sur d'autres groupes zoologiques comme les PorifŤres, ont rťvťlť des cas de plus en plus nombreux d'animaux appartenant ŗ la mÍme classe zoologique, quelquefois ŗ la mÍme famille, et possťdant des phosphagŤnes diffťrents.
La contribution la plus importante ŗ ces recherches a ťtť apportťe par Thoai et ses collaborateurs (Thoai, Roche, Robin et Thiem, 1955 ; Thoai et Robin, 1954; Roche et Robin. 1954 ; Thoai et Roche, 1957; Roche. Robin, Thoai et Pradel, 1960; Robin et Thoai. 1962); ils ont montrť "qu'aucun phosphagŤne ne caractťrise une classe ou une famille dťfinie d'animauxĽ (Thoai et Roche, 1957).
Un autre point ťlucidť au cours de ces dix derniŤres annťes (Thoai et Roche, 1964) est le rapport entre les phosphagŤnes et les guanidines libres dans les tissus d'un mÍme animal.
La base guanidique correspondant au phosphagŤne est, bien entendu, toujours prťsente, mais díautres composťs guanidiques qui ne participent pas ŗ la formation du phosphagŤne peuvent exister en grandes quantitťs chez un mÍme animal.
Tel est le cas de l'octopine chez les Cťphalopodes (Thoai, Roche et Robin. 1955 ; Thoai et Robin, 1959), de l'arcaÔne chez Arca noae (Roche, Robin, Thoai et Pradel. 1960), de la taurocyamine chez des PorifŤres (Roche et Robin. 1954 ; Robin et Roche, 1954).
D'autre part, il est presque de rŤgle que si un seul dťrivť guanidique est prťsent dans le muscle on le retrouve comme constituant du phosphagŤne musculaire.
Un autre cas intťressant est celui de la crťatine et de la phosphocrťatine chez des animaux comme Arenicola marina L., Leptosynapta inhaerens Muller, Martasterias glacialis L. (Roche, Thoai et Robin. 1957), Diadema setosum (Yanagisawa, 1959), oý ces corps sont prťsents seulement dans les spermatozoÔdes et non dans les autres cellules ou tissus de l'organisme.
L'ťtude d'un phosphagŤne ne devrait jamais Ítre sťparťe de celle des dťrivťs guanidiques prťsents dans les animaux d'oý le phosphagŤne a ťtť isolť; elle devrait toujours Ítre complťtťe par celle de la phosphotransfťrase correspondante.
Les deux premiers enzymes de ce groupe, la crťatine kinase (EC.2.7.3.2) et l'arginine kinase (EC.2.7.3.3) ont ťtť mises en ťvidence il y a trente ans par Lohmann (1954, 1955).
En 1957, Thoai a dťcrit la taurocyamine kinase et la glycocyamine kinase (EC.2.7.3.1) ; en 1960, Rosenberg et collaborateurs la lombricine kinase, et en 1962 Thoai, Robin et Pradel l'hypotaurocyamine kinase (1965).
Aussi est-ce en tenant compte de cet ensemble de points de vue que nous avons entrepris l'ťtude du phosphagŤne et de l'ATP : guanidine phosphotransfťrase d'Ophelia neglecta quand, il y a deux ans, nous avons remarquť la prťsence d'un dťrivť guanidique nouveau dans cette espŤce.
Pour la mÍme raison, nous avons ťtendu nos recherches ŗ un autre reprťsentant de la famille des Ophelidae.
OBJET DU TRAVAIL
Les recherches qui font l'objet du prťsent travail ont portť d'abord sur l'analyse chromatographique des guanidines monosubstituťes de deux espŤces de PolychŤtes sťdentaires appartenant au genre Ophelia, O. neglecta Schneider et O. bicornis Savigny.
Les dťrivťs guanidiques musculaires, qui entrent dans la constitution des phosphagŤnes, ont ťtť ensuite isolťs, cristallisťs et analysťs.
Nous avons montrť que les bases guanidiques musculaires, et donc les phosphagŤnes correspondants de deux espŤces animales voisines, sont diffťrents.
Chez O. bicornis elle est constituťe par la lombricine, qui n'a ťtť rencontrťe jusqu'ŗ prťsent que chez les OligochŤtes.
Chez O. neglacta, la base guanidique est un nouveau dťrivť monosubstituť, qui a ťtť identifiť et dont la structure a ťtť confirmťe par synthŤse. Ce corps a ťtť appelť ophťline.
Le phosphagŤne correspondant, la phosphoophťline, a ťtť ťgalement isolť et identifiť. Le produit de phosphorylation chimique de l'ophťline que nous avons prťparť peut servir de donateur de phosphate en prťsence d'ADP et d'une phosphotransfťrase extraite díO. neglecta.
Le nouvel enzyme, appelť ATP : ophťline phospho-transfťrase, qui catalyse la rťaction rťversible

a ťtť prťparť et ses propriťtťs examinťes.
L'ensemble des faits rapportťs dans le prťsent travail contribue en premier lieu ŗ la connaissance de nouveaux types de bases guanidiques et de phosphagŤnes.
En mettant en ťvidence le degrť de spťcificitť de la nouvelle ATP : guanidophosphotransfťrase et l'analogie de certaines de ses propriťtťs avec celles des enzymes du mÍme type, cette ťtude permet ťgalement des rapprochements utiles du point de vue de la biochimie comparťe.
RESUME
1į- un nouveau dťrivť guanidique a ťtť isolť du muscle d'Ophelia neglecta Schneider, purifiť sur colonne de rťsine ťchangeuse d'ions et obtenu ŗ l'ťtat cristallisť.
Le corps auquel on a donnť le nom díophťline a pour composition ťlťmentaire :
C= 24,4% ; H= 6,1% ; N= 21,3% ; P= 15,7%. Point de fusion : 272įC.
2į- Líophťline est une guanidine monosubstituťe rťpondant ŗ la formule du diester guanidoťthyl-mťthyl-phosphorique :
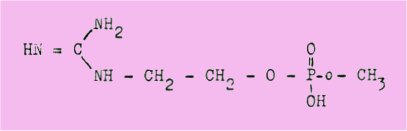
Sa constitution a ťtť ťtablie par identification des produits de son hydrolyse et par
comparaison du produit naturel isolť avec le mÍme composť prťparť par synthŤse chimique
(mÍme point de fusion, mÍme comportement chromatographique, mÍmes produits d'hydrolyse).
3į- Toutes les bases guanidiques prťsentes chez .O. neglecta ont ťtť dosťes dans trois types diffťrents de tissus.
4į- Le phosphagŤne d'O. neglecta, l'acide N'-phosphoryl-guanido-ťthyl-mťthyl-phosphorique, est lui aussi un corps inconnu jusqu'ŗ prťsent.
Le produit naturel a ťtť isolť, ses produits d'hydrolyse identifiťs et dosťs. La structure en a ťtť confirmťe par synthŤse et par comparaison des deux corps du point de vue des propriťtťs chimiques et de leurs comportements enzymatiques.
La prťsence d'un tautomŤre instable a ťtť discutťe.
5į- Nous avons trouvť dans le muscle d'O. neglecta une activitť enzymatique spťcifique catalysant le transfert d'un radical phosphoryl de l'ATP ŗ l'ophťline ou de la phosphophťline ŗ l'ADP selon la rťaction rťversible :
Nous avons extrait et purifiť cet enzyme que nous avons appelť ATP : ophťline phosphotransfťrase.
Les valeurs optima de pH et de tempťrature sont respectivement 8,5 et 30įC pour la rťaction directe (1) et 6,8 et 35įC pour la rťaction inverse (2).
Pour la rťaction directe, Vmax. = 31,6 mmoles par ml, par min. et par mg de protťine enzymatique, Km = 3 mM dans le cas de l'ATP et 5,8 mM dans le cas de l'ophťline.
Pour la rťaction inverse, Vmax = 19,6 mmoles par ml, par min. et par mg, Km = 0 ,5 mM dans le cas de l'ADP et 1,1 mM dans le cas de la phosphophťline.
6į- L'ATP : ophťline phosphotransfťrase est assez stable ŗ 0įC.
Elle est activťe par Mg++, inhibťe par le p-chloromercuribenzoate, la chloroacťtophťnone, la N-ťthylmalťimide, le monoiodoacťtate et l'hydroxylamine.
Elle est inactive sur la crťatine, l'arginine, la glycocyamine et les phosphagŤnes
correspondants; mais elle est active sur la lombricine, la taurocyamine et la
phosphotaurocyamine.
7į- Ni le nouveau phosphagŤne, ni la nouvelle base guanidique ne sont prťsents dans une autre espŤce du mÍme genre Ophelia examinťe, l'O. bicornis Savigny.
Chez celle-ci, une seule base guanidique musculaire est prťsente, la lombricine, qui n'a ťtť trouvťe jusqu'ŗ prťsent que chez des OligochŤtes, oý elle entre dans la constitution du phosphagŤne musculaire.
8į- La nouvelle mťthode d'isolement de la lombricine constitue une amťlioration apprťciable sur les techniques prťcťdemment utilisťes pour l'obtention de cette base ŗ partir des OligochŤtes.
La lombricine isolťe ŗ partir d'O. bicornis a ťtť cristallisťe et identifiťe et les bases guanidiques prťsentes chez ce polychŤte ont ťtť dosťes dans trois types diffťrents de tissus.
9)- Du point de vue de la biochimie de l'ťvolution, il est remarquable que deux espŤces d'un
mÍme genre, Ophelia, possŤdent deux guanidines musculaires diffťrentes ; ophťline pour O. neglecta et lombricine pour O. bicornis, celle-ci ťtant considťrťe jusqu'ŗ prťsent comme constituant caractťristique du phosphagŤne des OligochŤtes.
Le fait mťrite díautant plus d'Ítre soulignť qu'une autre espŤce de la mÍme famille d'Ophťlidťs, Travisia forbesii, possŤde dans le muscle deux types de phosphagŤnes diffťrents : phosphocrťatine et phosphoarginine, dont la coexistence est considťrťe jusqu'ici comme caractťristique des groupes zoologiques de transition (ťchinodermes).
MOTS CLEFS : ADP / animal / arginine / ATP / bicornis / biochimie / constitution / crťatine / dťrivť / enzyme / ťtude / guanidine / guanidique / kinase / lombricine / muscle / Needham / neglecta / oligochŤte / ophelia / ophťline / phosphagŤne / phosphagŤne / phosphoarginine / phosphocrťatine / phosphotransfťrase / rťaction / recherche / synthŤse / taurocyamine / tissus